Review Article - (2016) Volume 5, Issue 2
We report the creation of variant maps based on bat echolocation call recordings. The maps show regular patterns while characteristic features change when bat call recording properties change. By focusing on specific visual features, we found a set of projection parameters which allowed us to classify the variant maps into two distinct groups. These results are promising indicators that variant maps can be used as basis for new echolocation call classification algorithms.
Keywords: Echolocation; Algorithms; Morphometry; Fourier analysis; Quaternions.
The identification of echolocation calls is essential to the research and conservation of bat species [1]. However, automatic classification algorithms have not yet been proven capable of providing 100% correct classifications or getting closes enough to this ideal performance [2]. Since our approach of using variant maps shows already promising results, we are confident it will continue adding valuable contributions to the field of automatic bat call identification [3].
Automated bat echolocation call identification algorithms are developed since the end of 1990s [4-7]. Here, multivariate discriminant function analysis or neural networks were used for the classification of the calls. Since then, other methods were applied, e.g. algorithms of pattern recognition [8], support vector machines [9], hierarchical ensembles of neural networks [9,10], geometric morphometry [11], machine learning [12], CART [13] and random forest classification [14]. For a critical analysis of the performance of the applied methods we refer to Russo and Voigt [2] and the references therein.
Using variant maps for the classification of bat echolocation calls diers completely from these conventional techniques.
The main dierence is the pre-processing step, where the recordings are transformed into variant maps. This step oers the possibility to analyse the bat call recordings from a complete dierent point of view. It provides additional degrees of freedom which allow a further optimization of the identification process, e.g. by supplementing the information obtained from a Fourier analysis of the bat calls.
Our method to transform the bat call recordings is based on measures proposed by Zheng and Maeder [15] and Zheng [16] in the 1990s to partition special phase spaces in binary image analysis. These methods were extended in the 2010s [3,17] and successfully used to classify quantum interactions [18,19], dierently encrypted messages [20] and non-coding DNA [21-23].
Similar to these works, we transform the bat call recordings using variant measures to obtain variant maps. Each recording contains several calls of one bat species. There are four aerial-hawking bat species calling while flying across three types of crop fields far away from woody vegetation. The created variant maps have a regular structure, but characteristic features vary strongly with each recording. These results show that variant maps can be used to extract usable information from bat echolocation recordings.
The processed bat echolocation calls were recorded with a sampling rate of 500 kHz and saved as “raw” 16 bit audio files. In the following, we describe in four steps (A-D) how we transformed these files into variant maps.
Step A: From analogue to digital audio
In a recording of data length N, the amplitude of the bat echolocation calls is stored in N samples. Each sample corresponds to a floating point number of 16 bits. For simplicity, we transformed the floating point numbers to integer numbers of 16 bits.
Step B: From digital audio to quaternions
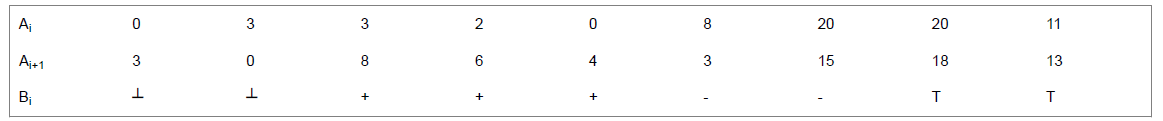
Next, we transform the integer sequence into a sequence of four meta states {┴, +, −, ┱} which resemble the quaternions {Bottom, Plus, Minus, Top}. For this step we select the i-th sample Ai and its next neighbor Ai+1 and define the dierence ΔA=Ai+1–Ai and local average L=(Ai+Ai+1)/2. Additionally we require the maximum Amax and minimum Amin of the current sequence to define a middle value V=(Amin+Amax)/2 and we define a tolerance T. Using these values we transform the integer sequence A1...AN into a sequence of quaternions B1...BN by using the rules:
if ΔA
if ΔA
if ΔA ≥ T and Ai>Ai+1 : Bi=−
if ΔA ≥ T and Ai
As an example, the values T=4 and V=10 lead to the sequence:
Step C: From quaternions to meta measures
We subdivide the quaternion sequence into segments of length M and obtain in this way S=N/M segments. For each segment, we define four meta measures {M┴, M+, M−, M┱}. One measure represents the number of associated quaternions in one segment. These meta measures satisfy the relations 0 ≤ M┴, M+, M−, M┱ ≤ M and M┴+M+ +M−+M┱=M. The quaternion sequence with N units is now represented by S segments where each segment contains four meta measures.
Step D: From meta measures to variant maps
There are many possibilities to combine meta measures for the creation of variant maps [3-23]. To transform the bat echolocation calls into 2D color maps we defined for each segment of meta measures the axis values X=M++M┴ and Y=M┴+M−+M┱. One Z value is obtained by counting the number of segments where one specific X-Y combination was found. Each Z value is represented by a color in an (M+1) × (M+1) matrix.
As an example, we depicted in Figure 1 the variant map of an echolocation call recording from the bat species Nyctalus noctula . It has a data length N=967139 and we chose a segment length M=237. At the position X=80 and Y=200 marked by a white circle, the color indicates a value Z=10. That is, we found 10 segments where the conditions M++M┴=80 and M┴+M−+M┱=200 apply. White areas indicate regions without any projection point on this sequence. For a discussion of further visual features which appear in this figure.
These type of maps oer the possibility to visualize long data sequences with >106 samples on compact matrices. We use this scheme to transform each bat call recording into a 2D color figure. It can be optimized for the identification of bat species, recording locations or times.

Figure 1: The variant map of an echolocation call recording from the species Nyctalus noctula created by following the processing steps A-D as described. We highlighted the position X=80 and Y=200 by a white circle to illustrate the processing step D. At this position the conditions M++M-=80 and M-+M-+MT=200 apply. Further visual features are as discussed in more detail.
Variant maps
Our main result is that all variant maps we created from bat echolocation calls show regular patterns while characteristic visual features vary with each recording. In the following, we describe the data we processed in detail and discuss the visual features we observed.
Data description
We processed 44 files which were recorded in August 2012 in the Uckermark region (Brandenburg, Germany) [24]. Each recording contains only calls of one of the four European bat species Nyctalus noctula , Pipistrellus nathusii , Pipistrellus pipistrellus or Pipistrellus pygmaeus . These files were recorded on arable fields cultivated with the three dierent crop types: corn (C), rapeseed (R) or wheat (W). The record length varies between 30 s to 2 min.

Figure 2: Variant maps of (a) Pipistrellus nathusii and (b) Nyctalus noctula , both recorded on a rapeseed field. The figures were created by applying the transformation process described in Figure (a) shows a typical double-maxima map with two significant maxima, while Figure (b) belongs to the group of non-double-maxima maps.
Visual features
We transformed all 44 files of bat calls into variant maps by processing the steps A to D as described. That is, we used the axis values X=M++M┴ and Y=M┴+M−+MT and a segment length M=237. By focusing on the visual features we clustered the resulting maps into two groups. A typical member of each group is shown in Figure 2.
One group consists only of maps showing patterns which have two significant maxima with values >105. We call members of this group double-maxima maps. The example shown in Figure 2(a) has maxima at the positions X=0, Y=237 and X=120, Y=200. Besides these two maxima, there are distinct positions on diagonal areas with values of the orders 1 to 103.
All other maps belong to the group of non-double-maxima maps. As an example, the map in Figure 2(b) has its significant maximum at the position X=0, Y=237, while other projection regions have values of the orders 1 to 103. In addition, most values of interest are located around a diagonal region and form a slat band on the map.
All 44 resulting maps are shown in Figures 3 and 4. They are separated into double-maxima maps (Figure 3) and non-doublemaxima maps (Figure 4). In principle, it is possible to further subdivide the variant maps by identifying additional visual features. However, since we did not yet find a direct connection between visual features and bat call properties, a further subdivision goes beyond the scope of this manuscript and will be the topic of a future publication.

Figure 3: These variant maps show double-maxima patterns. They have two significant maxima with values >105. The axis ranges are the same as in Figure 2. Each map origins from a bat echolocation recording on a corn (C), rapeseed (R) or wheat (W) field.

Figure 4: These variant maps show non-double-maxima patterns. That is, they explicitly do not have two distinct maxima with values >105 in contrast to the double-maxima maps shown in Figure 3.
On all generated maps, the positions on the left-down triangle area are empty. This is because our choice of axis obeys X+Y ≥ M. Empty positions in the right-upper area appear because the bat call recordings consist of discrete short pulses with a longer time period of silence in between.
Similarly, other visual characteristics in the colored areas can be directly related to properties of the bat call recordings. As an example, a signal of constant frequency can be transformed into a single position on a variant map by choosing suitable parameters. This means that, by optimizing the variant map transformation, it is possible to focus on features of the initial bat echolocation call for the creation of variant maps.
Since this transformation process is completely dierent from conventional bat call identification schemes, because it involves quaternion structures, it can be used to add optimizing parameters to these schemes and form in this way the basis for a new identification algorithm.
We transformed 44 bat echolocation files into variant maps. All created variant maps have a similar structure and can be classified by focusing on specific visual features. As an example, we found a set of projection parameters which allowed us to classify the recordings into double-maxima and non-double-maxima maps.
Features like this can be traced back to the signal nature of the recordings. In this way, variant maps oer the possibility to focus on individual features of bat echolocation calls. Since there are multiple numbers of possible combinations to create variant maps, we are very positive that a suitable projection combination can be found to fulfill our ultimate goal of identifying single bat species.
In order to meet this target it is necessary to process a much higher number of bat calls to create a sucient large database for the eective determination of possible projections and associated maps. This would form the perfect basis for the development of a new echolocation call identification algorithm.
We thank Voigt CC for providing the processed bat echolocation data and Troxell SA for revising the manuscript. Financial support by the National Science Foundation of China NSFC (No. 61362014) and the Overseas Higher-level Scholar Project of Yunnan Province, China, is gratefully acknowledged. Moreover, we appreciate the financial support by the Federal Ministry of Science, Research and Culture in Brandenburg, the University of Potsdam, the Leibniz Institute for Zoo and Wildlife Research and the Deutsche Forschungsgemeinschaft (Vo 890/22).
