Research Article - (2013) Volume 2, Issue 1
Failure to auto resuscitate from hypoxic-induced apnea by gasping has been suggested to play a role in sudden infant death. Little is known, however, about factors that influence gasping and its ability to sustain life during acute hypoxia in the newborn. Given that cardiac dysfunction and circulatory failure likely play a role in mediating auto resuscitation failure, the present experiments were carried out on 206 rat pups to investigate the influence of heart glycogen -- a metabolic substrate that supports cardiac function during acute hypoxia -- on the time to last gasp (TLG) during a single period of unrelenting hypoxia, and on the ability to auto resuscitate (AR) from primary apnea during repeated hypoxic challenge. On days 1-2, 5-6, and 10-11 postpartum, each pup was placed into a temperature controlled chamber regulated to 37 ± 1° C and exposed either to a single period of unrelenting hypoxia produced by breathing an anoxic gas mixture (97% N2 and 3% CO2) and the TLG determined, or repeatedly exposed to hypoxia and the ability to AR from primary apnea determined. Pups were studied with normal heart glycogen concentrations and with decreased heart glycogen concentrations effected by prior isoproterenol-induced transient tachycardia. A decrease in heart glycogen concentration did not significantly alter the TLG upon exposure to a single period of unrelenting hypoxia. It did, however, significantly decrease the number of successful AR from primary apnea during repeated hypoxic exposures in 1-2, 5-6, and 10-11 day-old pups. Thus, heart glycogen concentration influences the ability of rat pups to AR from hypoxic-induced primary apnea during early postnatal life.
Keywords: Apnea; Auto resuscitation; Gasping; Postnatal age; SIDS
Upon exposure to progressive and unrelenting hypoxia or asphyxia, newly born animals of a number of species display a sequential respiratory response which consists of hyperpnoea, primary apnea, gasping and secondary or terminal apnea [1-4]. Primary apnea may be as short as 10 seconds in newly born rats or as long as 18 minutes in newly born ground squirrels [5,6] and is often viewed as the interval between failure of the “centres” which control eupneic respiration and the resumption of respiratory activity in the form of gasping governed by medullary loci which at least in the rat are located in the lateral tegmental field [7]. Gasping following primary apnea occurs when the PaO2 decreases to ~8-10 torr; this is true during hyper capneic hypoxia produced by airway obstruction or during hypocapneic hypoxia produced by inhalation of a hypoxic gas mixture [2,3]. Peiper [8], Stevens [9], and Thach [10] have emphasized the importance of gasping in effecting “self-resuscitation” [5] or “auto resuscitation” [3] in human infants and that repeated exposure to hypoxia may lead to auto resuscitation failure and death.
Interestingly, we have found that postnatal age not only influences the number of hypoxic episodes tolerated by rat pups before auto resuscitation failure but also the sequence of events (i.e., mechanisms) leading to autoresuscitation failure [11]. For example, we have found that 5 to 6 day-old rat pups tolerate an average of 15 ± 2 hypoxicinduced apnea/auto resuscitation cycles before auto resuscitation failure whereas 10 to 11 day-old rats pups tolerate an average of only 11 ± 3 hypoxic-induced apnea/auto resuscitation cycles before auto resuscitation failure. Furthermore, during the terminal hypoxicinduced apnea/auto resuscitation cycle, the majority of 5 to 6 day-old pups display stages I (i.e., gasping) and II (i.e., cardiac resuscitation with a rapid increase in heart rate) of auto resuscitation occur before the onset of a trioventricular (AV) dissociation and the subsequent loss of ventricular depolarization; gasping continues throughout. This suggests that blood pressure is maintained during hypoxia and that gasping results in the transport of oxygen from the lungs to the heart with re oxygenation of a trial pacemaker cells. The subsequent arrhythmia which appears during the tachycardia of stage II of auto resuscitation may result from the cardiac effects of adenosine, the myocardial interstitial concentrations of which are well known to increase during hypoxia [12-15]. In 10 day and older pups, however, only stage I of auto resuscitation occurs before their demise. This suggests that blood pressure is not maintained during hypoxia and that gasping does not result in the transport of oxygen from the lungs to the heart. The inability of the heart to maintain cardiac output and blood pressure during hypoxia may result from the depletion of the cardiac metabolic substrate, glycogen during repeated exposure to hypoxia.
Our current experiments have been designed to investigate the role of heart glycogen in influencing protective responses of newly born rat pups to hypoxia. Specifically, we have determined a) basal heart glycogen concentration as well as changes following repeated exposure to hypoxia during early postnatal maturation, and b) heart glycogen concentration following isoproterenol-induced tachycardia during early postnatal maturation. In addition, we have carried out experiments to test the hypothesis that a decrease in heart glycogen concentration following isoproterenol-induced tachycardia alters the time to last gasp upon exposure to hypoxia as well as the ability of rat pups to auto resuscitate from hypoxic-induced primary apnea during early postnatal maturation.
Experiments were carried out on 208 Sprague-Dawley rat pups born by spontaneous vaginal delivery. Until an experiment, each pup was housed with its mother and siblings in a Plexiglas cage lined with Aspen-Chip Laboratory Bedding (North eastern Products) and kept in a humidity (30-40%) and temperature (25 ± 1°C, the selected ambient temperature and thermo neutral temperature of non pregnant and pregnant rat) controlled environmental chamber on a 12:12 h lightdark cycle with lights on from 0700. Although 25°C is below the thermo neutral zone of newborn rats [16], each pup had an opportunity to huddle with its siblings and dam in the nest, and thus to thermo regulate behaviourally.
All experimental procedures described herein were carried out in accordance with the “Guide to the Care and Use of Experimental Animals” provided by the Canadian Council on Animal Care, and with the approval of the Animal Care Committee of the University of Calgary.
Experimental protocols
Experimental Series I – Heart Glycogen and Plasma Glucose following Multiple Hypoxic Induced Apnea/Auto resuscitation Cycles (HIA/AR) during Postnatal Maturation: For an experiment, each 1 to 2 (n=23), 5 to 6 (n=24) and 10 to 11 (n=23) day-old rat pup was removed from its mother and siblings, weighed and instrumented for measurement of cardiovascular and respiratory variables. The pup was positioned prone in a metabolic chamber regulated to 37.0 ± 0.1°C into which flowed room air at a rate of 1 litre per minute. After 30 minutes, the pup was exposed to 0, 1, 5, or 10 HIA/AR at 5 minute intervals before it was removed from the chamber and rapidly decapitated for collection of trunk blood for determination of plasma glucose concentration and removal of the heart for determination of glycogen content. For each HIA/AR, the gas which flowed into the chamber was changed from room air to 97% N2 and 3% CO2 until primary apnea occurred; the gas was then changed back to room air and auto resuscitation was effected by gasping. When the gas mixture was changed, the flow rate was increased until the gas concentrations in the chamber had stabilized; the flow rate was then lowered to 1 litre per minute. This procedure resulted in equilibration and stabilization of the new gas mixture in approximately 5 seconds. An upper limit of 10 HIA/AR was chosen as we have previously found that naïve 10 to 11 day-old pups studied at a thermo neutral temperature of 37.0°C tolerate an average of 11 episodes of HIA/AR before auto resuscitation failure [11]. During an experiment, stages of the respiratory response to hypoxia were directly observed on the polygraph tracing.
Experimental Series II – Heart Glycogen and Plasma Glucose after Isoproterenol Induced Tachycardia during Postnatal Maturation: For an experiment, each 1 to 2 (n=17), 5 to 6 (n=16) and 10 to 11 (n=15) day-old rat pup was removed from its mother and siblings, weighed and instrumented for measurement of cardiovascular and respiratory variables. The pup was positioned prone in a metabolic chamber regulated to 37.0 ± 0.1°C into which flowed room air at a rate of 1 litre per minute. After 30 minutes, the pup was removed from the chamber and given a subcutaneous injection of 1.25 mg of isoproterenol per kg of body weight; the pup was then returned to the metabolic chamber. At 120 minutes, the pup was removed from the chamber and rapidly decapitated for collection of trunk blood for determination of plasma glucose concentration and removal of the heart for determination of glycogen content.
The rationale for using 1.25 mg/kg isoproterenol in Experimental Series II was as follows. In preliminary dose – response experiments carried out on 1 to 2, 5 to 6 and 10 to 11 day-old rat pups and testing doses of 0, 0.312, 0.625, 1.25, 2.5, 5.0, 10.0 mg/kg of the non selective beta adrenergic receptor agonist isoproterenol, we determined 1.25 mg/ kg to be the smallest dose that elicited a maximal heart rate response (i.e., the EC100) in all age groups and in which heart rate returned back to control levels by ~120 minutes.
Experimental Series III – Time to Last Gasp during a Single Unrelenting Anoxic Gas Challenge after Isoproterenol Induced Tachycardia during Postnatal Maturation: For an experiment, each 1 to 2 (n=22), 5 to 6 (n=16) and 10 to 11 (n=18) day-old rat pup was removed from its mother and siblings and instrumented for measurement of cardiovascular and respiratory variables. The pup was then positioned prone in a metabolic chamber regulated to 37.0 ± 0.1°C into which flowed room air at a rate of 1 litre per minute for 30 minutes before it was removed briefly from the chamber and given a subcutaneous injection of vehicle or 1.25 mg/kg isoproterenol. After another 120 minutes, the pup was exposed to a period of unrelenting hypoxia by changing the gas which flowed into the metabolic chamber from room air to 97% N2 and 3% CO2 and the gasping and heart rate responses were determined on a minute-to-minute basis until the onset of secondary or terminal apnea.
Experiment IV – Autoresuscitation From Primary Apnea during Repeated Anoxic Gas Challenges after Isoproterenol Induced Tachycardia during Postnatal Maturation: For an experiment, each 1 to 2 (n=10), 5 to 6 (n=12) and 10 to 11 (n=12) day-old rat pup was removed from its mother and siblings and instrumented for measurement of cardiovascular and respiratory variables. The pup was then positioned prone in a metabolic chamber regulated to 37.0 ± 0.1°C into which flowed room air at a rate of 1 litre per minute for 30 minutes before it was removed briefly from the chamber and given a subcutaneous injection of vehicle or 1.25 mg/kg isoproterenol. After another 120 minutes, the gas which flowed into the metabolic chamber was changed from room air to 97% N2 and 3% CO2 until primary apnea occurred; the gas was then changed back to room air and the ability of the pup to auto resuscitate by gasping determined; this procedure was repeated at five-minute intervals until auto resuscitation failure occurred. During an experiment, primary apnea was detected by directly observing respiratory movements on the polygraph tracing.
In order to determine if auto resuscitation had occurred, we counted heart rate and respiratory rate during the last 15 seconds of each five minute period. If heart rate and respiratory rate were greater than 60% of control values, we deemed the auto resuscitation a success. If, however, heart rate and/or respiratory rate during the last 15 seconds of each five minute period were less than 60% of control values, we deemed the auto resuscitation a failure.
Experimental apparatus
The metabolic chamber used in our experiments consisted of a double-walled plexiglass cylinder (30 cm long - internal diameter 6 cm) into which flowed room air or 97% N2 and 3% CO2.Chamber ambient temperature was regulated to 37.0 ± 0.1°C by circulating water from a temperature controlled bath (Neslab - Endocal Refrigerated Circulating Bath RTE-8DD) through the space between the walls. The rationale for studying the various age groups of rats at an ambient temperature of 37.0°C was as follows: Our laboratory has previously determined the selected ambient temperatures of 1 to 2, 5 to 6 and 10 to 11 day old rats, 20 to 30 minutes after being placed into a 200 cm thermocline with a linear temperature gradient of 25°C to 40°C to be as follows [11]: 1 to 2 days - 36 ± 1°C; 5 to 6 days - 37 ± 1°C; 10 to 11 days - 37 ± 2°C (mean ± 1SD). Numerous experiments carried out on newborn as well as older animals provide evidence that when given the opportunity, animals select an ambient temperature that is within their thermo neutral zone [16-21].
Experimental measurements and calculations
During an experiment, the electrocardiogram, respiratory movements and chamber O2 and CO2 levels were recorded on a Model 7 polygraph (Grass Instrument Company) at a paper speed of 10 mm sec-1. Leads I, II and III of the electrocardiogram were recorded from multi stranded stainless steel wire electrodes (AS 633, Cooner Wire Company) sewn on the right shoulder, left shoulder and the left thigh; the electrodes were connected to Model 7HIP5 High Impedance Probes coupled to a Model 7P5 Wide Band EEG A.C. Preamplifiers (Grass Instrument Company). Respiratory movements were recorded from a mercury-in-silicone rubber strain gauge (Model HgPC, D.M. Davis, Inc.) placed around the chest; the strain gauge was connected to a bridge amplifier (Biomedical Technical Support Center, University of Calgary) that was coupled to a Model 7P03 Adapter Panel (Grass Instrument Company). Chamber oxygen and carbon dioxide levels were measured using an Applied Electrochemistry Carbon Dioxide and Oxygen Analyzer (Ametek) coupled to a Model 7P03 Adapter Panel.
Glucose
Following decapitation, blood was collected and glucose measured immediately by an Accu-check compact plus blood sugar meter.
Glycogen
Following decapitation, the heart was rapidly excised and placed into liquid nitrogen and then stored at -80°C until glycogen was measured using a Glycogen Assay Kit (Biovision, Catalog #K646-100). In this assay, gluco amylase hydrolyzes the glycogen to glucose which is then specifically oxidized to produce a product that reacts with Oxi Red probe to generate color (λmax = 570 nm) and fluorescence (Ex 535/Em587).
Statistical analysis was carried out by ANOVA or ANOVA for repeated measures where appropriate and followed by a Holm-Sidak multiple comparison test. All results are reported as means ± one standard deviation, and p<0.05 was considered to be of statistical significance.
Basal heart glycogen concentration decreased with increasing postnatal age and averaged 4.1 ± 0.7 mg per gram of heart tissue on postnatal days 1 to 2, 3.5 ± 0.5 mg per gram of heart tissue on postnatal days 5 to 6, and 3.0 ± 0.5 on postnatal days 10 to 11 (p=0.012). Plasma glucose concentration, on the other hand, increased with increasing postnatal age and averaged 4.6 ± 0.6 mmol per liter of blood on postnatal days 1 to 2, 5.9 ± 0.4 mmol per litre of blood on postnatal days 5 to 6, and 6.6 ± 0.4 mmol per litre of blood on postnatal days 10 to 11 (p<0.001).
Experimental Series I – Heart Glycogen and Plasma Glucose following Multiple HIA/AR Cycles during Postnatal Maturation: Exposure to multiple HIA/AR cycles at 5-minute intervals produced a gradual decrease in heart glycogen which was significant at 5 HIA/AR cycles in 1 to 2, 5 to 6 and 10 to 11 day old pups (Figure 1) (ANOVA - HIA/AR <0.001; Age <0.001; HIA/AR × Age 0.763). After 10 HIA/ AR cycles, heart glycogen decreased 58% in 1 to 2 day old pups, 63% in 5 to 6 day old pups, and 62% in 10 to 11 day old pups as comparedto control. To the contrary, plasma glucose increased in 1 to 2 and 5 to 6 day old pups and stayed the same in 10 to 11 day old pups following exposure to multiple HAI/AR cycles (Figure 2) (ANOVA - HIA/AR <0.001; Age <0.001; HIA/AR × Age 0.175).
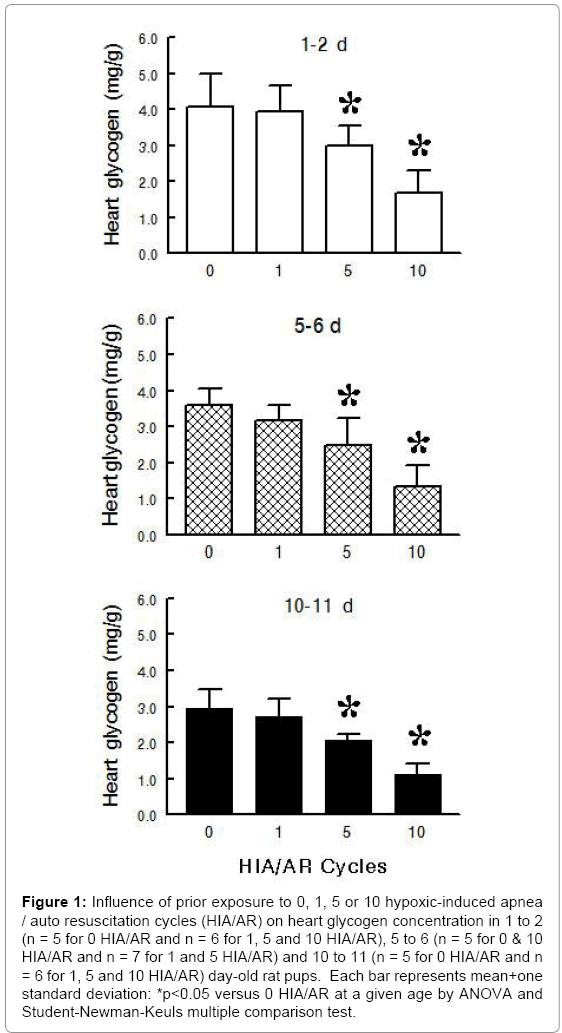
Figure 1: Influence of prior exposure to 0, 1, 5 or 10 hypoxic-induced apnea / auto resuscitation cycles (HIA/AR) on heart glycogen concentration in 1 to 2 (n = 5 for 0 HIA/AR and n = 6 for 1, 5 and 10 HIA/AR), 5 to 6 (n = 5 for 0 & 10 HIA/AR and n = 7 for 1 and 5 HIA/AR) and 10 to 11 (n = 5 for 0 HIA/AR and n = 6 for 1, 5 and 10 HIA/AR) day-old rat pups. Each bar represents mean+one standard deviation: *p<0.05 versus 0 HIA/AR at a given age by ANOVA and Student-Newman-Keuls multiple comparison test.
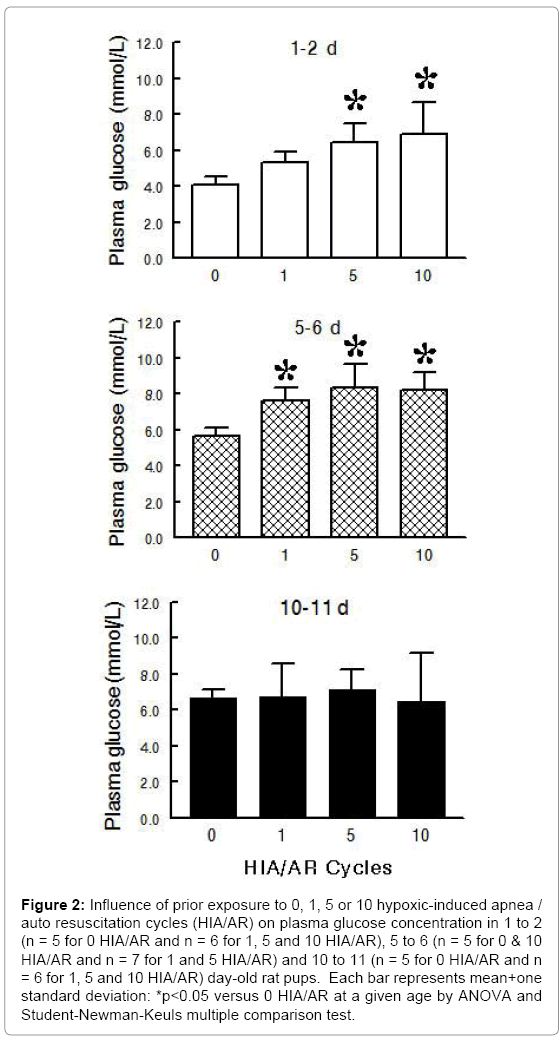
Figure 2: Influence of prior exposure to 0, 1, 5 or 10 hypoxic-induced apnea / auto resuscitation cycles (HIA/AR) on plasma glucose concentration in 1 to 2 (n = 5 for 0 HIA/AR and n = 6 for 1, 5 and 10 HIA/AR), 5 to 6 (n = 5 for 0 & 10 HIA/AR and n = 7 for 1 and 5 HIA/AR) and 10 to 11 (n = 5 for 0 HIA/AR and n = 6 for 1, 5 and 10 HIA/AR) day-old rat pups. Each bar represents mean+one standard deviation: *p<0.05 versus 0 HIA/AR at a given age by ANOVA and Student-Newman-Keuls multiple comparison test.
Experimental Series II – Heart Glycogen and Plasma Glucose following Isoproterenol Induced Tachycardia during Postnatal Maturation: Heart glycogen decreased after isoproterenol induced tachycardia in all age groups (Figure 3) (ANOVA - Drug <0.001; Age 0.003; Drug × Age 0.612) but plasma glucose was unaffected (Figure 4) (ANOVA - Drug 0.578; Age <0.001; Drug × Age 0.869).After isoproterenol induced tachycardia, heart glycogen decreased 58% in 1 to 2 day old pups, 43% in 5 to 6 day old pups, and 50% in 10 to 11 day old pups as compared to saline.
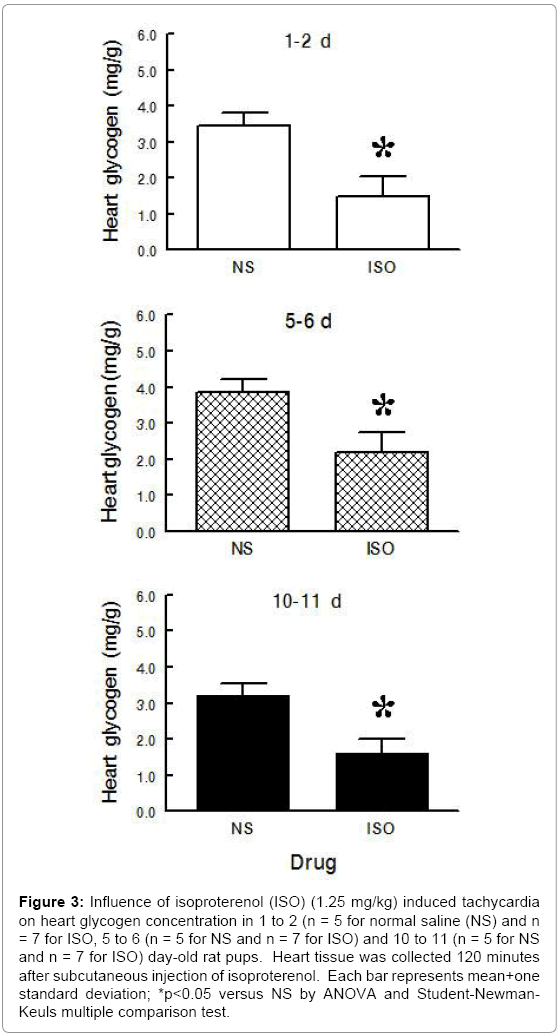
Figure 3: Influence of isoproterenol (ISO) (1.25 mg/kg) induced tachycardia on heart glycogen concentration in 1 to 2 (n = 5 for normal saline (NS) and n = 7 for ISO, 5 to 6 (n = 5 for NS and n = 7 for ISO) and 10 to 11 (n = 5 for NS and n = 7 for ISO) day-old rat pups. Heart tissue was collected 120 minutes after subcutaneous injection of isoproterenol. Each bar represents mean+one standard deviation; *p<0.05 versus NS by ANOVA and Student-Newman- Keuls multiple comparison test.
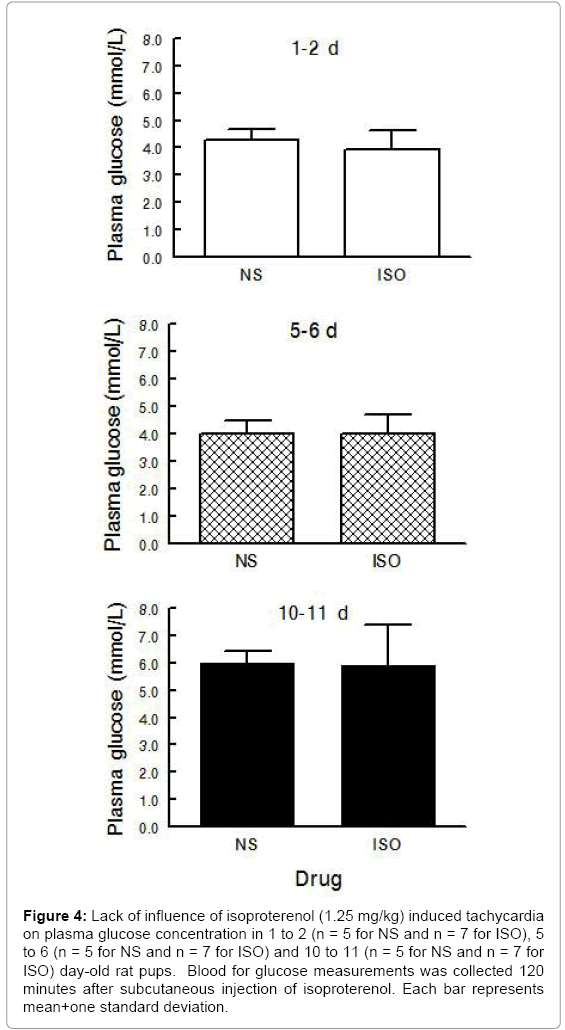
Figure 4: Lack of influence of isoproterenol (1.25 mg/kg) induced tachycardia on plasma glucose concentration in 1 to 2 (n = 5 for NS and n = 7 for ISO), 5 to 6 (n = 5 for NS and n = 7 for ISO) and 10 to 11 (n = 5 for NS and n = 7 for ISO) day-old rat pups. Blood for glucose measurements was collected 120 minutes after subcutaneous injection of isoproterenol. Each bar represents mean+one standard deviation.
Experimental Series III – Time to Last Gasp during a Single Unrelenting Anoxic Gas Challenge after Isoproterenol Induced Tachycardia during Postnatal Maturation: The time to last gasp (ANOVA - Drug <0.224; Age <0.001; Drug × Age 0.249) in response to a single anoxic gas challenge was not significantly influenced by isoproterenol induced tachycardia and the resulting decrease in heart glycogen concentration (Figure 5). Heart rate decreased significantly during anoxic gas challenge (Figure 6); the heart rate response, however, was not influenced by prior exposure to isoproterenol and the associated tachycardia or decrease in heart glycogen concentration.
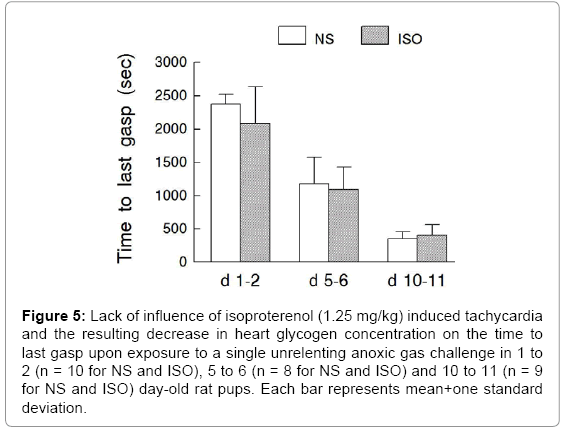
Figure 5: Lack of influence of isoproterenol (1.25 mg/kg) induced tachycardia and the resulting decrease in heart glycogen concentration on the time to last gasp upon exposure to a single unrelenting anoxic gas challenge in 1 to 2 (n = 10 for NS and ISO), 5 to 6 (n = 8 for NS and ISO) and 10 to 11 (n = 9 for NS and ISO) day-old rat pups. Each bar represents mean+one standard deviation.
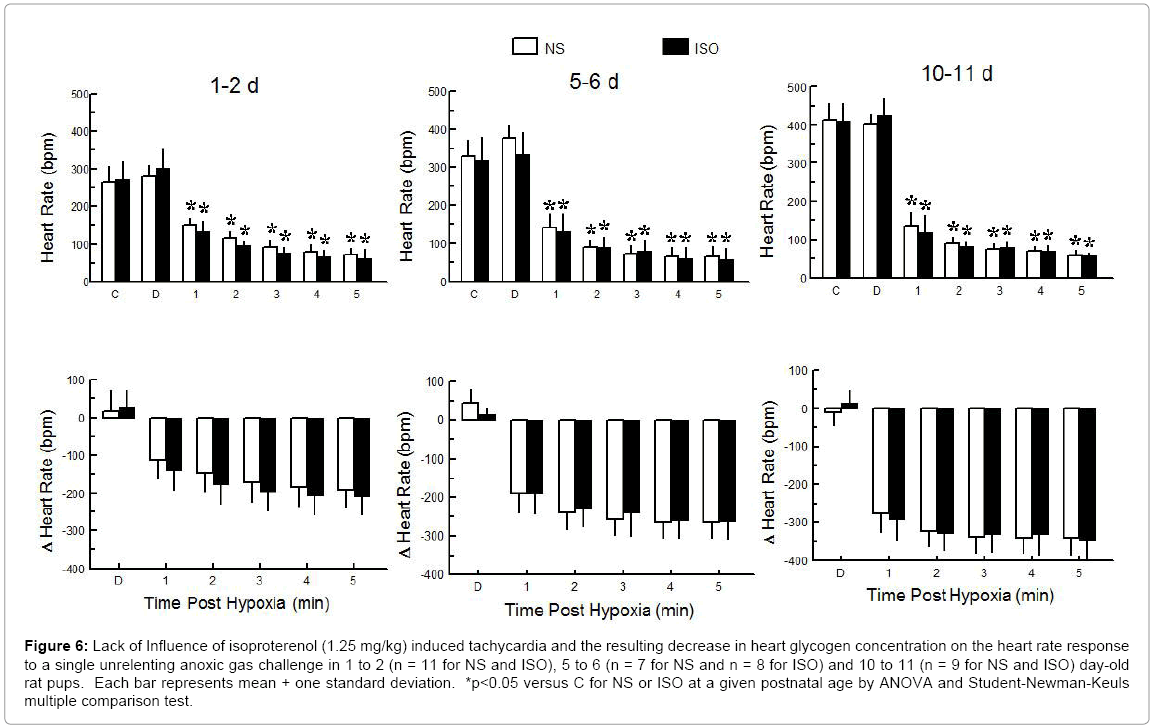
Figure 6: Lack of Influence of isoproterenol (1.25 mg/kg) induced tachycardia and the resulting decrease in heart glycogen concentration on the heart rate response to a single unrelenting anoxic gas challenge in 1 to 2 (n = 11 for NS and ISO), 5 to 6 (n = 7 for NS and n = 8 for ISO) and 10 to 11 (n = 9 for NS and ISO) day-old rat pups. Each bar represents mean + one standard deviation. *p<0.05 versus C for NS or ISO at a given postnatal age by ANOVA and Student-Newman-Keuls multiple comparison test.
Experiment IV – Auto resuscitation From Primary Apnea during Repeated Anoxic Gas Challenges after Isoproterenol Induced Tachycardia during Postnatal Maturation: The number of successful auto resuscitations from primary apnea during repeated exposure to anoxic gas challenge decreased significantly following isoproterenol induced tachycardia and the resulting decrease in heart glycogen concentration in 1 to 2, 5 to 6, and 10 to 11 day-old pups (Figure 7) (ANOVA - Drug 0.007; Age <0.001; Drug × Age 0.445). The decrease in the number of successful auto resuscitations following isoproterenol was relatively constant when expressed as a percent change being -31% in 1 to 2 day-old pups, -32% in 5 to 6 day-old pups, and -27% in 10 to 11 day-old pups compared to that observed following saline.
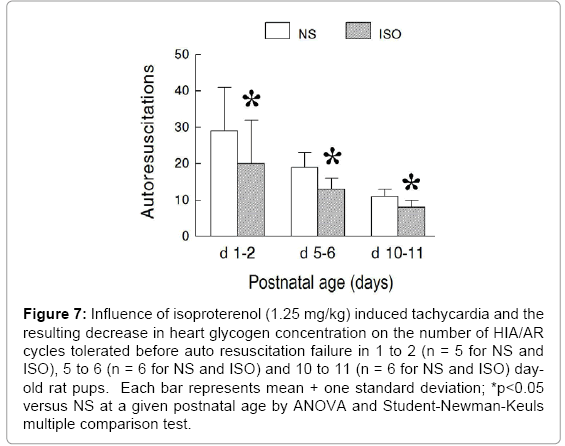
Figure 7: Influence of isoproterenol (1.25 mg/kg) induced tachycardia and the resulting decrease in heart glycogen concentration on the number of HIA/AR cycles tolerated before auto resuscitation failure in 1 to 2 (n = 5 for NS and ISO), 5 to 6 (n = 6 for NS and ISO) and 10 to 11 (n = 6 for NS and ISO) dayold rat pups. Each bar represents mean + one standard deviation; *p<0.05 versus NS at a given postnatal age by ANOVA and Student-Newman-Keuls multiple comparison test.
Our experiments provide new information about heart glycogen and its role in supporting processes that promote survival of naïve rat pups upon exposure to hypoxia during early postnatal maturation. Novel findings in our study were that: a) basal heart glycogen concentration decreased and plasma glucose concentration increased during early postnatal maturation; b) repeated exposure to hypoxia produced a gradual decrease in heart glycogen concentration but not plasma glucose concentration; c) transient isoproterenol-induced tachycardia produced a decrease in heart glycogen concentration but not plasma glucose concentration; d) transient isoproterenol-induced tachycardia and the resulting decrease in heart glycogen concentration did not alter the subsequent heart rate response to hypoxia; e) transient isoproterenol-induced tachycardia and the resulting decrease in heart glycogen concentration did not significantly alter the time to last gasp upon exposure to a single unrelenting anoxic gas challenge; and f) transient isoproterenol-induced tachycardia and the resulting decrease in heart glycogen concentration significantly decreased the number of successful autoresuscitations from primary apnea during repeated anoxic gas challenges. Thus, although heart glycogen concentration does not influence the duration of potential autoresuscitation producing gasps in rat pups upon exposure to a single period of unrelenting hypoxia, it does influence their ability to successfully autoresuscitate from repeated hypoxic-induced primary apneas during early postnatal life.
We found that basal heart glycogen concentration was highest in newly born rat pups and that it decreased during early postnatal maturation which is in keeping with the work of others including Dawes et al. [22], Stafford and Weatherall [23] and Shelley [24]. When oxygen demand exceeds oxygen supply such as in hypoxic hypoxia induced by anoxic gas challenge, there is a transition from aerobic to anaerobic metabolism; early experiments established that glycolysis provides essential energy for anaerobic metabolism and thus it has been suggested to promote survival during hypoxia in foetal, newborn and young animals [22,25,26]. For example, Dawes et al. [22] showed that survival of fatal sheep during hypoxia produced by umbilical cord ligation was related to basal carbohydrate concentration in the heart and reasoned accordingly that one of the limiting factors in anaerobic survival is the ability of the heart to maintain the circulation; survival in their experiments was assessed by the maintenance of blood pressure and heart rate. Hypoxia does elicits gasping in the foetus [3,27] but this maneuver is futile with regard to re oxygenation given the status of the lungs and pulmonary circulation and the fluid-filled environment of the uterus which surrounds the foetus; ultimately, foetal survival as such is dependent upon reestablishment of umbilical blood flow and placental oxygenation. The situation is quite different in the newborn as successful auto resuscitation from hypoxic-induced apnea -- such as may occur during sleep related obstructive apnea or positional asphyxia -- depends upon reoxygenation of the blood by the respiratory system via gasping as well as transport of the oxygenated blood to the heart and brain by the cardiovascular system [28].
With regard to the newborn, Dawes et al. [22] have also pointed out that a direct correlation exists between basal carbohydrate concentration in the hearts of newborn rats, rabbits and guinea pigs and survival during hypoxia produced by anoxic gas challenge; in these experiments, however, survival was assessed by the respiratory variable time to last gasp, and not the maintenance of blood pressure and heart rate as was used to assess survival in the foetus. Although the close correlation between cardiac glycogen concentration and the time to last gasp during anoxia in the newborn was emphasized, it is questionable that this represents cause and effect as we have found that the time to last gasp during exposure to anoxia and the time to last gasp following decapitation which arrests the circulation, are similar in 5 to 6 day-old rat pups studied in a thermo neutral environment [29]. The relatively tight correlation between heart carbohydrate concentration and time to last gasp observed in the intact animal could be due to the short and consistent interval of time between failure of the respiratory system and cardiovascular system during hypoxia as has been shown in dogs and puppies [30,31]. In the present experiments, we found that a reduction in heart glycogen concentration to about 50% of normal following transient isoproterenol-induced tachycardia did not significantly influence the time to last gasp; a result perhaps not surprising given the results of our aforementioned decapitation experiments. A decrease in heart glycogen concentration did, however, influence the ability of rat pups to auto resuscitate from repeated exposure to hypoxic-induced apnea during early postnatal life, a manoeuvre that requires functional contribution and integration of several organs such as the brain, lungs, and adrenal glands.
Ten to 11 day-old rat pups studied in a thermo neutral environment tolerate an average of 11 hypoxic induced apnea/auto resuscitations cycles before failure [11]. In our current study, we found that cardiac glycogen concentration in 10 to 11 day-old rat pups decreased from a basal value of 2.9 ± 0.5 mg per gram of heart tissue to 1.1 ± 0.3 mg per gram of heart tissue after 10 hypoxic-induced apnea/auto resuscitation cycles (i.e., the hypoxic challenge prior to anticipated auto resuscitation failure). When cardiac glycogen concentration was decreased to a similar level following isoproterenol-induced transient tachycardia (i.e., 1.6 ± 0.4 mg per gram of heart tissue), the pups tolerated fewer hypoxic-induced apnea/auto resuscitation cycles before auto resuscitation failure suggesting that a critical level of cardiac glycogen is needed to support heart function and effect successful auto resuscitation in these pups; the mechanism of auto resuscitation failure (i.e., failure of gasping to effect stage II of auto resuscitation) was not altered. Our data therefore support the hypothesis that depletion of heart glycogen participates in mediating circulatory failure and auto resuscitation failure in 10 to 11 day old rat pups following repetitive exposure to hypoxia.
Not unexpectedly, we also found that decreasing heart glycogen concentration via isoproterenol-induced transient tachycardia decreased the number of hypoxic-induced apnea/auto resuscitation cycles before auto resuscitation failure in 1 to 2 and 5 to 6 day-old pups. In about half of these pups, a lack of gasping to effect stage II of auto resuscitation resulted in auto resuscitation failure as in the 10 to 11 dayold pups. The others displayed stages I (i.e., gasping) and II (i.e., cardiac resuscitation with a rapid increase in heart rate) of auto resuscitation before the onset of atrio-ventricular dissociation and the subsequent loss of ventricular depolarization. Perhaps in these animals with higher basal heart glycogen concentrations, the accumulation of adenosine mediated failure via atrio-ventricular dissociation before glycogen decreased below a level critical required supporting of mechanical activity of the heart during hypoxia. This postulate is currently being investigated in our laboratory.
Our results add to the literature and to those of Deshpande et al. [32] who measured cardiac glycogen concentration in BALB/c mice before and after repeated hypoxic-induced apnea/auto resuscitations cycles; their experiments were carried out on naïve 21 day-old BALB/c mice that normally tolerate an average of 6 hypoxic induced apnea/ auto resuscitations cycles before auto resuscitation failure. Upon their first exposure to hypoxia, these mice displayed a rapid decrease in heart glycogen concentration and interestingly exhibited no further measureable depletion with subsequent hypoxic induced apnea/auto resuscitations cycles. Because heart glycogen levels stabilized after the first hypoxic induced apnea/auto resuscitations cycle, the investigators concluded that a case could not be made for depletion of heart glycogen being a major factor in this type of auto resuscitation failure in older rodents. The difference between their results and ours may be related to species but most likely is related to a postnatal age effect and the fact that basal cardiac glycogen concentration decreases significantly during the first three weeks of postnatal life.
In conclusion, previous experiments from our laboratory have shown that a number of factors including postnatal age [11], core temperature [33], prior exposure to hypoxia [34], fetal exposure to nicotine [6,35,36], gender [37], and body position [38] significantly influence the number of anoxic challenges tolerated before auto resuscitation failure. Our present experiments extend these findings and provide evidence that heart glycogen promotes survival of naïve rat pups upon exposure repeated bouts of hypoxia during early postnatal life.
This study was supported by the Canadian Institutes of Health Research.
